夏枯草来源外泌体装载奥利司他通过调控脂代谢抑制肝癌细胞生长


肝细胞癌是全球常见且预后较差的恶性肿瘤,其治疗面临术后高复发与晚期耐药等挑战[1-3]。近年研究发现,肿瘤代谢重编程尤其是脂质代谢异常在肝癌发生发展中起关键作用[4-5]。肝癌细胞高度依赖内源性脂肪酸合成,其中脂肪酸合成酶(fatty acid synthase,FASN)作为限速酶显著高表达,成为潜在治疗靶点[6-9]。奥利司他是一种FASN抑制剂,具有抗肿瘤潜力,但其较强的疏水性导致水溶性极差,体内代谢迅速,生物利用度低,难以在肿瘤部位维持有效治疗浓度,限制了其临床转化与应用[10-12]。因此,寻找能够有效递送奥利司他的新型载体,改善其药动学特性,是实现其抗肿瘤价值的关键。外泌体作为天然纳米载体,具有良好的生物相容性与低免疫原性,植物来源外泌体更具备来源广泛、成本低廉等优势[13]。近年来,植物来源的外泌体因其具有生物相容性好、免疫原性低、稳定性高、易于跨越生物屏障以及来源广泛、成本低廉等优势,作为新型药物递送载体受到了广泛关注[14-15]。夏枯草具有抗肿瘤、抗炎和免疫调节等多种活性[16],推测源自夏枯草的外泌体不仅本身可能继承其母体植物的部分生物活性,更可作为奥利司他的理想天然纳米载体,构建一种基于天然产物的生物递送系统。因此,本研究拟构建夏枯草来源外泌体装载奥利司他(Prunella vulgaris-derived exosomes loading orlistat,PVENs-Orl)递送系统,探讨其对肝癌细胞脂代谢及恶性生物学行为的影响及潜在机制,为基于天然外泌体的脂代谢靶向治疗提供实验依据。
1 材料
1.1 细胞
人肝癌HepG2细胞(货号CL-0103)购自武汉普诺赛生命科技有限公司。
1.2 药材
新鲜夏枯草全株来源于湖南省中西医结合医院药房,经湖南省中西医结合医院曾普华教授及邓芳教授鉴定为唇形科植物夏枯草Prunella vulgaris L.的干燥全草。
1.3 药品与试剂
1.4 仪器
Optima XE型超速离心机(美国贝克曼库尔特有限公司);Q700型超声细胞破碎仪(美国Branson公司);HT7800型透射电子显微镜(transmission electron microscopy,TEM,日本日立有限公司);ZetaView型纳米颗粒追踪分析仪(nanoparticle tracking analysis,NTA,德国Particle Metrix公司);373528型徕卡显微镜(德国徕卡公司);Synergy H1型多功能酶标仪(美国BioTek公司);Heracell VIOS型细胞培养箱(美国Thermo Fisher Scientific公司);LightCycler®480实时荧光定量PCR仪(瑞士Roche公司)。
2 方法
2.1 PVENs的提取与表征
将新鲜夏枯草全株和磷酸盐缓冲液(phosphate buffered saline,PBS)溶液按1∶2的比例放入破壁器中,高速切碎5 min,并用纱布滤过。得到的汁液分别经1 000 r/min离心10 min、3 000 r/min离心20 min、10 000 r/min离心40 min,去除大的植物组织和细胞碎片。收集上清液,150 000 r/min超速离心2 h,并在PBS中重新悬浮。将悬浮液分别转移到15%、30%、45%和60%蔗糖中,150 000 r/min超速离心2 h,得到纯化的PVENs。
取10 μL PVENs悬液,滴于铜网,2%磷钨酸负染后,TEM下观察其形态结构。另取适量样品稀释后,通过NTA检测其粒径分布。
2.2 奥利司他的装载与PVENs-Orl的制备
采用孵育-超声法进行载药。将1 mg奥利司他溶于适量二甲基亚砜(dimethyl sulfoxide,DMSO)中,与1 mL含10 mg PVENs的PBS悬液混合,使DMSO终体积分数低于0.5%。将混合液于37 ℃摇床孵育2 h,随后置于冰浴中,使用超声细胞破碎仪(功率200 W,超声2 s,间隔3 s,总时长2 min)进行辅助载药。载药后的悬液再次经110 000×g超速离心70 min,以去除未包封的奥利司他,沉淀用PBS重悬,即得PVENs-Orl。通过NTA检测载药前后纳米颗粒的粒径与Zeta电位变化,以初步验证载药成功。
2.3细胞培养
HepG2细胞用含10%胎牛血清、100 U/mL青霉素和100 μg/mL链霉素的DMEM高糖培养基,置于37 ℃、5% CO2的饱和湿度培养箱中培养,取对数生长期的细胞用于后续实验。
2.4 CCK-8法检测细胞活力
将对数生长期的HepG2细胞以5×103个/孔接种于96孔板中,培养24 h待细胞贴壁后,更换为含有不同质量浓度(0、10、20、30、40、50 μg/mL)PVENs或PVENs-Orl的新鲜培养基,分别培养24、48、72 h。另设置不接种细胞不给药的空白组。每孔加入10 μL CCK-8溶液,孵育2 h后,用酶标仪测定450 nm处的吸光度(A)值,计算细胞存活率,并利用GraphPad Prism 9.0软件计算半数抑制浓度(half inhibitory concentration,IC50)。
细胞存活率=(A给药-A空白)/(A对照-A空白)
2.5 细胞摄取实验
采用绿色荧光探针DiO标记PVENs及PVENs-Orl。将DiO工作液与PVENs或PVENs-Orl悬液混合,37 ℃避光孵育30 min,110 000×g超速离心70 min去除游离DiO,PBS重悬沉淀,即得DiO-PVENs和DiO-PVENs-Orl。将对数生长期的HepG2细胞以2×105个/孔接种于共聚焦培养皿中,培养24 h贴壁后,分别加入等量的DiO-PVENs和DiO-PVENs-Orl,37 ℃避光共孵育4 h。弃去上清,PBS洗涤3次,4%多聚甲醛固定15 min,DAPI染核后,于共聚焦显微镜下观察并拍照。
2.6 细胞功能学实验
设置对照组、PVENs(1/2 IC50、IC50、2 IC50)组及PVENs-Orl(1/2 IC50、IC50、2 IC50)组和阳性对照索拉非尼(10 μmol/L)组,考察PVENs-Orl对HepG2细胞增殖、迁移和侵袭的影响。
2.6.1克隆形成实验将对数生长期的HepG2细胞以500个/孔接种于6孔板中,给药干预24 h后更换为新鲜完全培养基,继续培养10~14 d,结晶紫染色后计数大于50个细胞的克隆。
2.6.2划痕实验细胞长满单层后,用200μL枪头制造划痕,PBS清洗后更换为无血清培养基并给予相应药物干预,于0、24 h在显微镜下拍照,计算划痕愈合率。
划痕愈合率=(0 h划痕面积-24 h划痕面积)/0 h划痕面积
2.6.3Transwell侵袭实验在Transwell上室铺覆Matrigel基质胶,接种含2×104个细胞的无血清培养基,下室加入含10% FBS的培养基作为趋化剂,给药干预24 h后,擦去上室未侵袭细胞,下室细胞经4%多聚甲醛固定、结晶紫染色后,于显微镜下随机选取5个视野计数。
2.7 脂代谢表型分析
设置对照组、PVENs(IC50)组及PVENs-Orl(IC50)组和索拉非尼(10 μmol/L)组,给药干预后,考察PVENs-Orl对HepG2细胞关键脂代谢指标的影响。
2.7.1 FFA摄取采用CheKineTMFFA含量检测试剂盒,通过铜离子结合法,于550 nm波长下检测细胞中FFA含量。
2.7.2 FASN活性使用FAS活性检测试剂盒,通过检测340 nm处还原型烟酰胺腺嘌呤二核苷酸磷酸(nicotinamide adenine dinucleotide phosphate,NADPH)的消耗速率,反映FASN酶活性。
2.7.3 TG含量采用GPO-PAP法TG测试盒,于500 nm波长下测定细胞内TG含量。
2.8 脂质过氧化与细胞毒性检测
设置对照组、PVENs(IC50)组及PVENs-Orl(IC50)组和RSL(1 μmol/L)组,给药干预后,按照试剂盒说明书操作,加入DCFH-DA荧光探针,采用流式细胞术检测细胞内ROS水平;通过硫代巴比妥酸(thiobarbituric acid,TBA)比色法测定MDA水平;收集细胞上清液,按试剂盒说明书检测LDH活性,反映细胞膜的完整性及损伤程度。
2.9线粒体功能检测
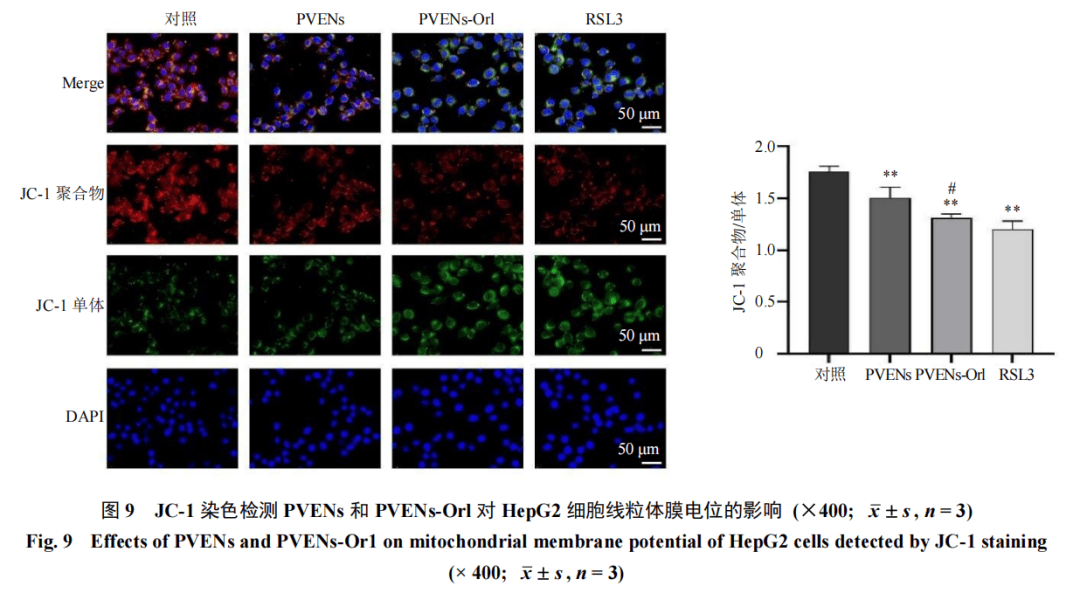
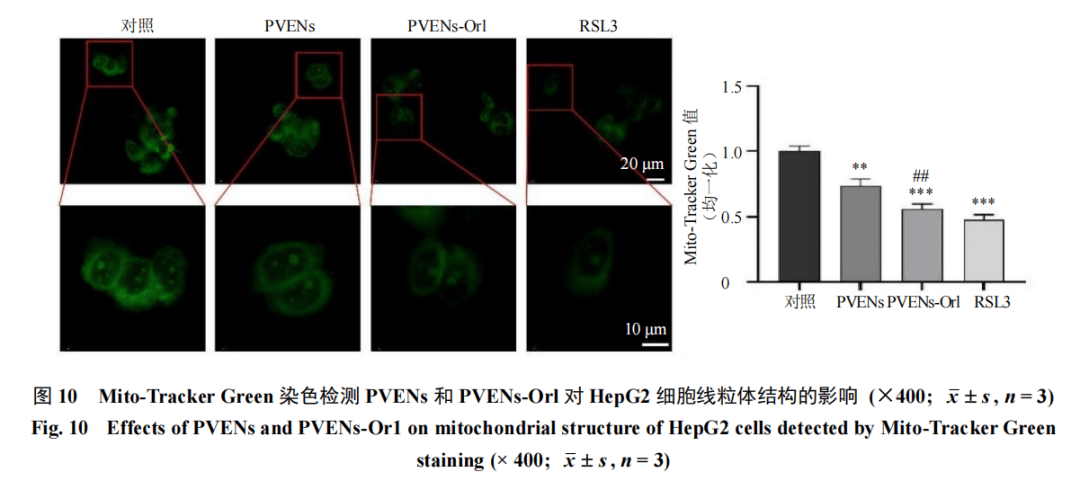
设置对照组、PVENs(IC50)组及PVENs-Orl(IC50)组和RSL(1 μmol/L)组,给药干预后,按照试剂盒说明书操作,加入JC-1荧光探针,于荧光显微镜下观察并拍照,以正常线粒体(红色荧光)与去极化线粒体(绿色荧光)的比例变化反映线粒体膜电位的高低。给药干预后,按照试剂盒说明书操作,加入Mito-Tracker Green探针,于荧光显微镜下观察线粒体结构并拍照,统计线粒体绿色荧光强度。
2.10 qRT-PCR检测脂代谢与铁死亡相关基因表达
设置对照组、PVENs(IC50)组及PVENs-Orl(IC50)组和RSL(1 μmol/L)组,给药干预后,使用TRIzol法提取细胞总RNA,测定浓度与纯度后,逆转录合成cDNA。以cDNA为模板,使用SYBR Green法在QuantStudio 5系统上进行qRT-PCR。以GAPDH为内参,采用2−∆∆Ct法计算各基因的相对表达量。

2.11统计学分析
采用GraphPad Prism 9.0软件进行统计分析,多组间比较采用单因素方差分析(One-way ANOVA),2组间比较采用Student’s t检验。数据以表示。
3结果
3.1PVENs的提取、表征及奥利司他装载
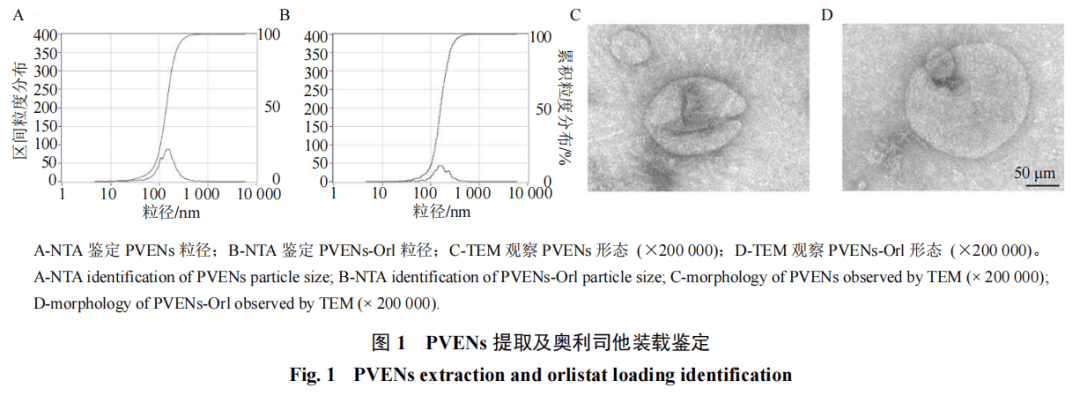
采用超速离心法成功从夏枯草水提物中分离出PVENs。TEM结果(图1-C)显示,PVENs呈典型的杯状或球形双层膜结构,形态完整。NTA结果(图1-A)表明,超高速离心法提取的PVENs粒径为(154.1±76.2)nm,粒径占比为100.0%。装载奥利司他后,所得PVENs-Orl的粒径与PVENs相比形态结构未发生显著变化,但具备更大的粒径(图1-B、D),表明其生物学特性仍得到保留。
3.2 HepG2细胞摄取PVENs结果
为验证PVENs及其载药体系能否作为有效的药物递送载体被靶细胞摄取,采用绿色荧光探针DiO分别对PVENs与PVENs-Orl进行膜标记。将等量的DiO-PVENs与DiO-PVENs-Orl分别与HepG2细胞共孵育后,通过共聚焦显微镜观察。如图2所示,与对照组相比,DiO-PVENs组和DiO-PVENs-Orl组细胞质内均出现了显著的绿色荧光信号。表明奥利司他的装载并未改变PVENs本身被HepG2细胞内化的能力,无论是PVENs还是PVENs-Orl均能有效地被HepG2细胞摄取,为其作为靶向递送系统并发挥后续生物学功能提供了直接证据。

3.3 PVENs-Orl抑制HepG2细胞增殖
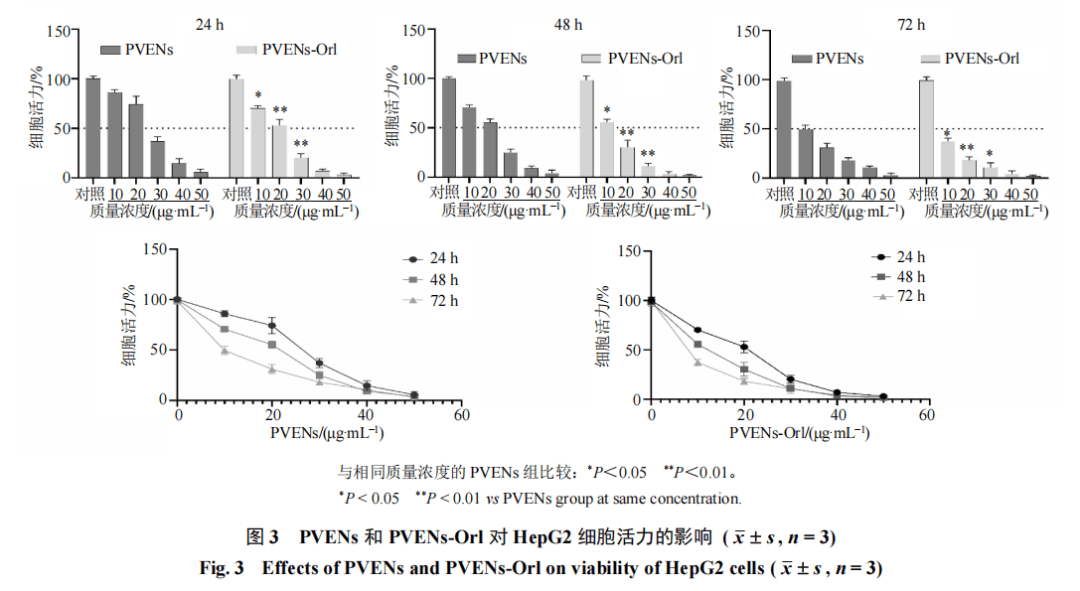
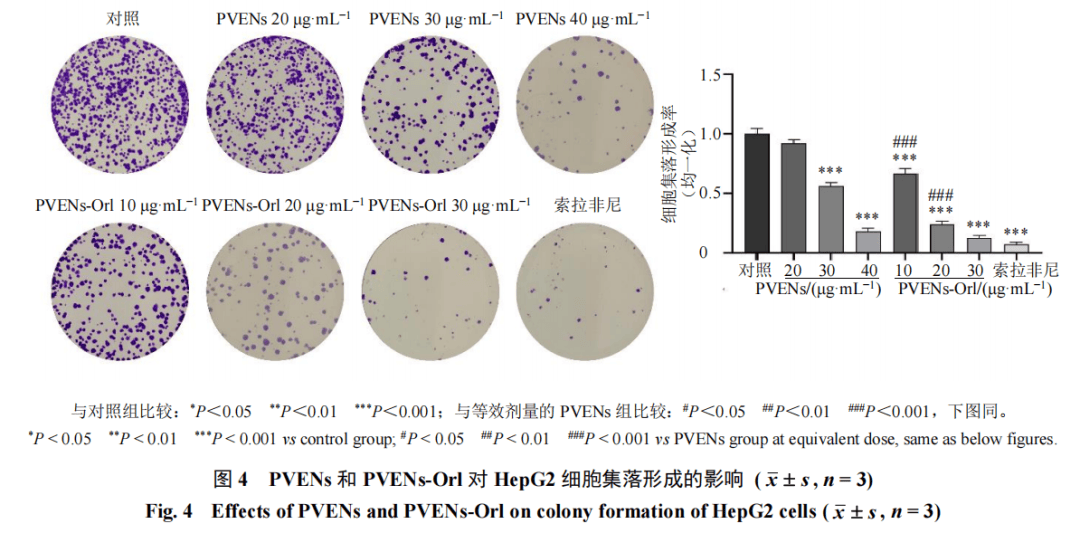
如图3所示,PVENs和PVENs-Orl均能够抑制HepG2细胞增殖,且呈剂量和时间相关性。对相同质量浓度、相同干预时间下PVENs组与PVENs-Orl组的细胞活性进行组间比较,结果显示,干预24、48、72 h,PVENs-Orl(10、20、30 μg/mL)组的细胞活性均显著低于PVENs组(P<0.05、0.01)。PVENs干预HepG2细胞24、48、72 h的IC50分别为25.0、23.0、10.0μg/mL,PVENs-Orl干预HepG2细胞24、48、72 h的IC50分别为22.0、13.0、4.0μg/mL。根据48 h的IC50值,设置1/2 IC50、IC50、2 IC50用于后续实验。细胞增殖、迁移及侵袭实验均采用等效药物浓度配对设计,PVENs-Orl(10、20、30μg/mL)为外泌体联合等量奥利司他,与PVENs(20、30、40 μg/mL)为等效暴露剂量,故进行组间比较。如图4所示,与对照组比较,PVENs(30、40 μg/mL)组及PVENs-Orl(10、20、30 μg/mL)组和索拉非尼组细胞集落大小、数量均显著减少(P<0.001),且PVENs-Orl组的细胞集落大小、数量明显小于PVENs组(P<0.001)。
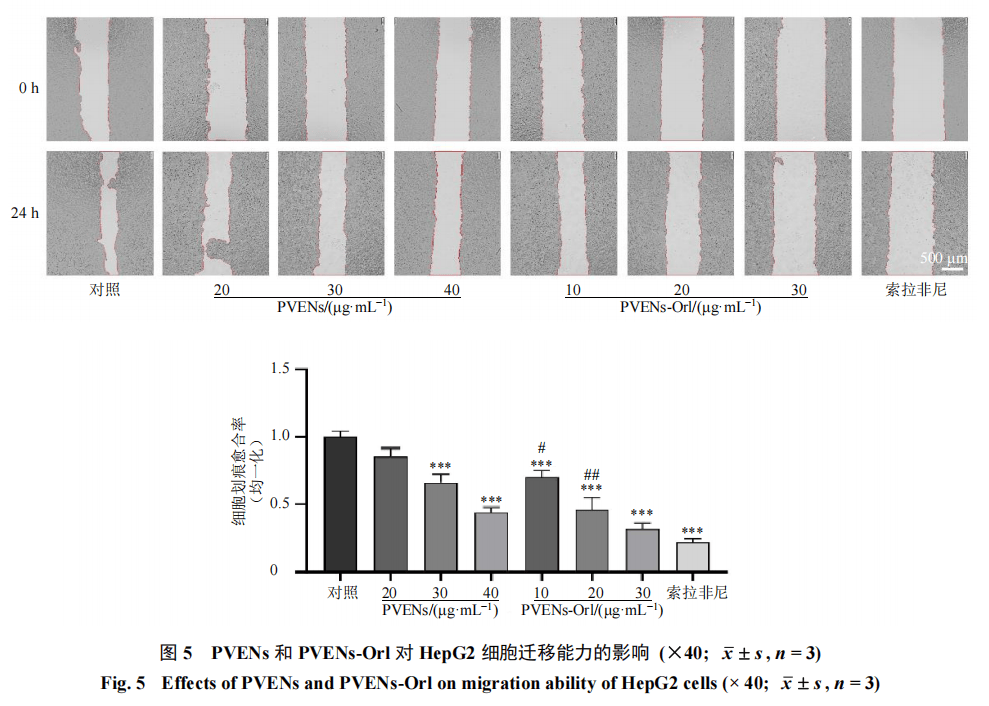
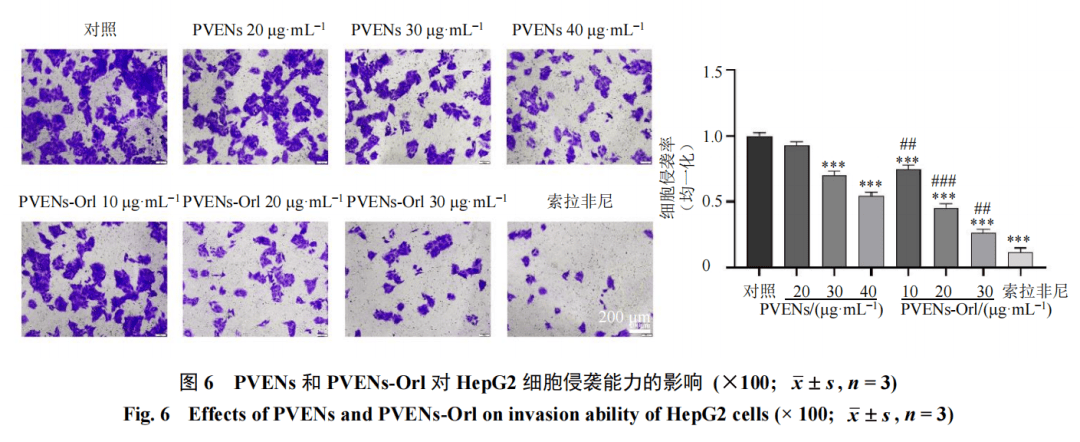
3.4 PVENs-Orl对HepG2细胞迁移和侵袭的影响
如图5所示,与对照组比较,PVENs(30、40 μg/mL)组及PVENs-Orl(10、20、30 μg/mL)组和索拉非尼组细胞划痕愈合率均显著降低(P<0.001);且等效剂量条件下,PVENs-Orl组的细胞划痕愈合率明显低于PVENs组(P<0.05、0.01),提示PVENs-Orl对HepG2细胞迁移能力的抑制作用强于PVENs。Transwell实验进一步验证了上述结果,如图6所示,对照组细胞穿膜率较高,而PVENs(30、40 μg/mL)组及PVENs-Orl(10、20、30 μg/mL)组和索拉非尼组细胞Transwell穿膜率均显著降低(P<0.001);其中PVENs-Orl组的细胞穿膜率显著低于PVENs组(P<0.01、0.001)。以上结果表明,PVENs可抑制HepG2细胞的迁移与侵袭能力,而装载奥利司他后的PVENs对HepG2细胞迁移、侵袭的抑制作用进一步增强。
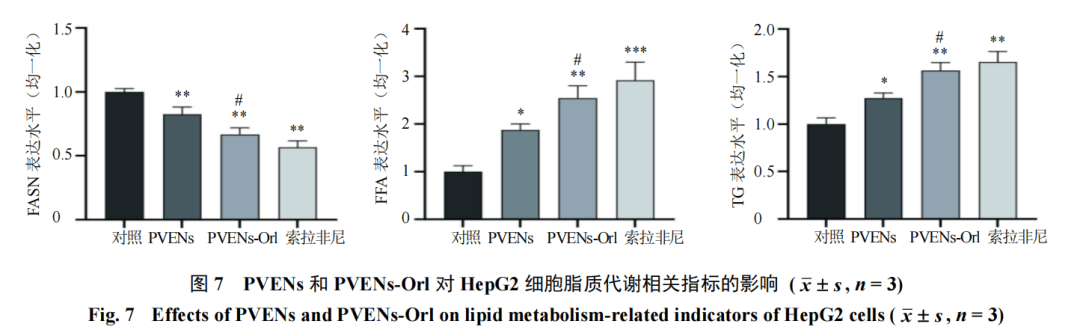
3.5 PVENs-Orl调控HepG2细胞的脂代谢表型
为探究PVENs-Orl抑制HepG2细胞生长的作用机制是否与调控脂代谢相关,检测了FFA摄取、FASN活性及TG含量等关键脂代谢指标。如图7所示,与对照组比较,PVENs组、PVENs-Orl组及索拉非尼组FASN水平均显著降低(P<0.01),而脂质代谢中间产物FFA及终产物TG的水平则明显升高(P<0.05、0.01、0.001)。其中PVENs-Or1组的调控效应较PVENs组更为显著(P<0.05),提示PVENs装载奥利司他可通过抑制脂质合成关键酶FASN的表达、促进脂质代谢产物FFA与TG的积累,实现对HepG2细胞脂代谢过程的重编程。
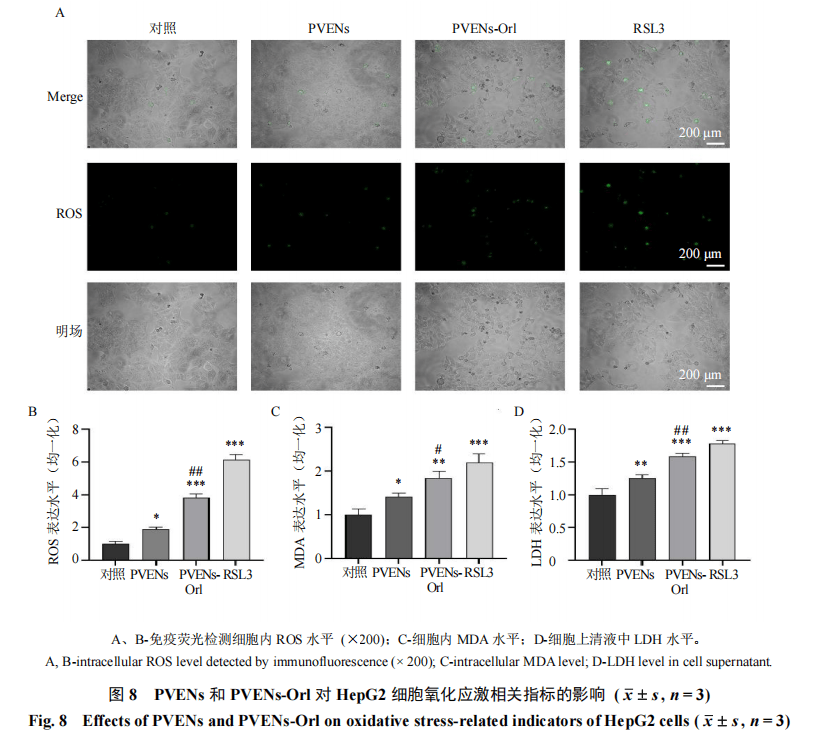
3.6 PVENs-Orl干预HepG2细胞脂质过氧化
采用DCFH-DA荧光探针检测细胞内ROS水平,结果如图8-A、B所示,与对照组比较,PVENs组、PVENs-Orl组及铁死亡诱导剂RSL3组细胞内ROS水平均显著升高(P<0.05、0.001)。如图8-C、D所示,与对照组比较,PVENs组、PVENs-Orl组及RSL3组细胞内MDA和上清液中LDH水平均显著升高(P<0.05、0.01、0.001)。且PVENs-Or1组的调控效应较PVENs组更为显著(P<0.05、0.01),表明PVENs-Orl引发了HepG2细胞严重的氧化应激,导致细胞膜完整性受损。
3.7 PVENs-Orl干预HepG2细胞脂质过氧化与线粒体相关
鉴于线粒体是细胞代谢和ROS产生的核心细胞器,而脂质过氧化与线粒体功能障碍存在紧密的相互调控关系,进一步检测了线粒体功能。JC-1染色结果(图9)显示,与对照组比较,PVENs组、PVENs-Orl组及RSL3组红色荧光(JC-1聚合物,代表正常高膜电位)强度显著减弱,绿色荧光(JC-1单体,代表去极化低膜电位)强度增强,且JC-1聚合物相对值显著下降(P<0.01),表明线粒体膜电位降低,而线粒体膜电位的下降会进一步促进ROS的产生与累积,加剧脂质过氧化进程。Mito-Tracker Green染色结果(图10)进一步证实,与对照组比较,PVENs组、PVENs-Orl组及RSL3组线粒体荧光强度显著降低(P<0.01、0.001),提示线粒体膜电位下降、功能受损。以上结果表明,PVENs-Orl能有效诱导HepG2细胞线粒体功能障碍。
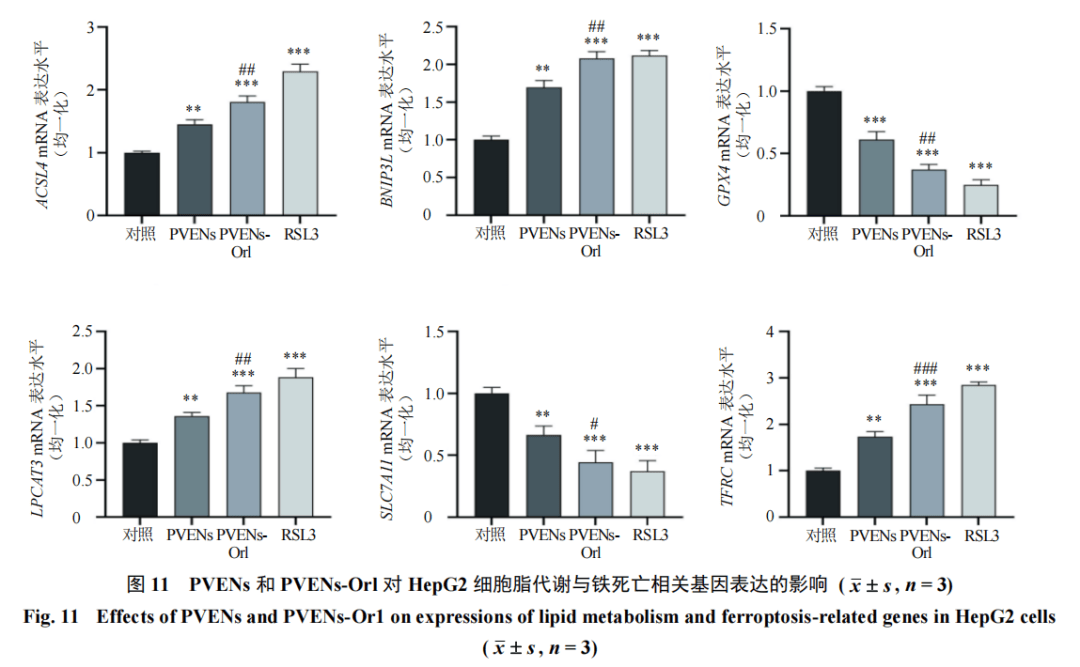
3.8PVENs-Orl对HepG2细胞脂代谢与铁死亡相关基因表达的影响
为进一步阐明PVENs-Or1调控HepG2细胞脂代谢的分子机制,采用qRT-PCR检测了铁死亡及脂代谢相关基因表达。如图11所示,与对照组比较,PVENs组、PVENs-Or1组及RSL3组脂质过氧化及铁死亡促进相关基因(ACSL4、BNIP3L、LPCAT3、TFRC)表达水平均显著升高(P<0.01、0.001),抗氧化及铁死亡抑制相关基因(GPX4、SLC7A11)表达水平均显著降低(P<0.01、0.001),且PVENs-Or1组的调控效应更接近铁死亡诱导剂RSL3组。以上结果表明,PVENs-Or1可通过上调促脂质过氧化/铁死亡基因、下调抗铁死亡基因的表达,调控脂代谢与铁死亡的协同通路,进而抑制HepG2细胞生长。